
ΠΡΟΤΕΙΝΟΜΕΝΟ ΘΕΜΑ Δ, για πανελλαδικές 2023
Στο παρακάτω γενεαλογικό δέντρο απεικονίζεται μια κληρονομική μονογονιδιακή ασθένεια του μεταβολισμού, σε μια οικογένεια. ...
Read More

ΘΕΜΑ Α
Α1 – δ
Α2 – γ
Α3 – β
Α4 – β
Α5 – β
ΘΕΜΑ Β
Β1: Α – 3, Β – 2, Γ – 3, Δ – 2, Ε – 1, ΣΤ – 1, Ζ – 1
όδον αφορά την επιλογή Δ: ένα μόριο tRNA μπορεί να είναι συζευγμένο με αμινοξύ. Όμως, στην επιλογή 3 αναγράφεται “αμινοξέα” και δεν μπορεί να αντιστοιχηθεί με το tRNA
Β2: Παραμέτσιουμ, μυικά κύτταρα, ώριμα ερυθρά αιμοσφαίρια.
Β3:
Το ερώτημα που αντιμετωπίσαμε στην αρχή του κεφαλαίου ήταν: «Πώς είναι δυνατό να έχουμε τα ίδια βασικά ανθρώπινα χαρακτηριστικά με τα συγγενικά μας πρόσωπα, χωρίς να είμαστε πανομοιότυποι μεταξύ μας;». Τώρα που γνωρίζουμε τις λεπτομέρειες της μείωσης ίσως μπορούμε να το απαντήσουμε. Η μείωση σε συνδυασμό με τη γονιμοποίηση διασφαλίζει στο δημιουργούμενο ζυγωτό μια πλήρη διπλοειδή σειρά χρωμοσωμάτων και γονιδίων. Έτσι ο οργανισμός που θα προέλθει από αυτό εκδηλώνει, όπως οι γονείς του και τα αδέλφια του, το σύνολο των βασικών γνωρισμάτων που προσδιορίζει το είδος τους. Ταυτόχρονα όμως κάθε οργανισμός έχει πάρει από τους γονείς του, μέσω των γαμετών τους, μια συλλογή χρωμοσωμάτων και γονιδίων, που είναι απίθανο να υπάρχει σε κάποιο από τα αδέλφια του. Αυτή η μοναδική συλλογή αποκτάται, όπως είδαμε, χάρη στους δύο μηχανισμούς, τον ανεξάρτητο συνδυασμό χρωμοσωμάτων και τον επιχιασμό. Χάρη στον ανεξάρτητο συνδυασμό χρωμοσωμάτων δημιουργείται ένα πλήθος από νέους συνδυασμούς μη ομόλογων χρωμοσωμάτων και συνεπώς ένα πλήθος από νέους συνδυασμούς γονιδίων, που βρίσκονται σε μη ομόλογα χρωμοσώματα. Η απλοειδής σειρά χρωμοσωμάτων συμβολίζεται με n. Η διπλοειδής, αντίστοιχα, συμβολίζεται με 2n. Στον άνθρωπο για παράδειγμα, n=23 και 2n=46. Όταν ένα κύτταρο με 2n χρωμοσώματα υφίσταται μείωση για την παραγωγή γαμετών, τότε οι διαφορετικοί συνδυασμοί μη ομόλογων χρωμοσωμάτων που μπορούν να εμφανιστούν σε διαφορετικούς γαμέτες (απλοειδή n κύτταρα) που θα προκύψουν από αυτήν είναι 2n. Αυτό για τον άνθρωπο σημαίνει ότι κάθε γονέας έχει καταθέσει σε κάθε γαμέτη του τον έναν από τους 223 συνδυασμούς που μπορεί να παραγάγει. Σε αντίθεση με τον ανεξάρτητο συνδυασμό χρωμοσωμάτων, ο οποίος έχει ως αποτέλεσμα την αναδιανομή των γονιδίων που βρίσκονται σε μη ομόλογα χρωμοσώματα, ο επιχιασμός ανασυνδυάζει γονίδια που βρίσκονται στο ίδιο το ζεύγος ομόλογων χρωμοσωμάτων. Αυτό συμβαίνει, γιατί με την ανταλλαγή αντίστοιχων τμημάτων, που γίνεται μεταξύ των μη αδελφών χρωματίδων των ομόλογων χρωμοσωμάτων, ανταλλάσσονται και γονίδια. Ο συνδυασμός των δύο μηχανισμών που αναφέρθηκαν έχει ως συνέπεια σε κάθε γαμέτη να αντιπροσωπεύεται ένα μοναδικό «μείγμα» γονιδίων που βρίσκονται σε διαφορετικά χρωμοσώματα και ταυτόχρονα ένα μοναδικό «μείγμα» γονιδίων που βρίσκονται στο ίδιο χρωμόσωμα. Έτσι λοιπόν, χάρη στη μείωση, είναι στατιστικά απίθανο εμείς και κάποιο από τα αδέλφια μας να έχουμε την ίδια συλλογή χρωμοσωμάτων και γονιδίων και από τους δύο γονείς, οπότε να είμαστε πανομοιότυποι μεταξύ μας. Το γεγονός αυτό, που είναι η ουσία της γενετικής ποικιλομορφίας που χαρακτηρίζει τους αμφιγονικά αναπαραγόμενους οργανισμούς, έχει μεγάλη σημασία για την εξέλιξη. Μερικοί από τους συνδυασμούς γονιδίων (άρα και γνωρισμάτων που επηρεάζονται από τα γονίδια αυτά) είναι επιτυχέστεροι απ’ ό,τι άλλοι, με την έννοια ότι προσφέρουν μεγαλύτερες δυνατότητες επιβίωσης στο φορέα τους σε συγκεκριμένες περιβαλλοντικές συνθήκες. Ο μηχανισμός αυτός συμβάλλει στην εξέλιξη, γιατί κάθε πληθυσμός περνά στις επόμενες γενιές του πιο ευνοϊκούς συνδυασμούς γονιδίων και γνωρισμάτων.
Β4: α) Μείωση
β)Μετάφαση – 2
γ) i) 6 χρωμοσώματα / 12μόρια DNA
ii) 3 χρωμοσώματα / 3 μόρια DNA
ΘΕΜΑ Γ
Γ1:
Το αντικωδικόνιο του tRNA που μεταφέρει το αμινοξύ τρυπτοφάνη έχει αλληλουχία βάσειων 3’ACC5′, καθώς είναι συμπληρωματικό και αντιπαράλληλο του κωδικονίου mRNA 5’UGG3′.
Η μεταγραφόμενη αλυσίδα γονιδίου (μη κωδική) είναι συμπληρωματική και αντιπαράλληλη με το παραγόμενο mRNA. Ισχύον οι κανόνες συμπληρωματικότητας των βάσεων: Α (αδενίνη) στη μη κωδική συμπληρωματική με U (ουρακίλη) στο RNA, Τ (θυμίνη) στη μη κωδική συμπληρωματική με Α στο RNA, G (γουανίνη) συμπληρωματική με C (κυτοσίνη).
Η μη μεταγραφόμενη αλυσίδα γονιδίου (κωδική) είναι συμπληρωματική και αντιπαράλληλη με την μη κωδική. Κατ’ επέκταση, κωδική αλυσίδα και RNA που συντίθεται από τη μεταγραφή του γονιδίου είναι ίδιας αλληλουχίας με τη διαφορά πως αντί U που εντοπίζεται στο RNA υπάρχει Τ στην κωδική του γονιδίου.
Με βάση αυτά, στην κωδική αλυσίδα για το γονίδιο που μεταγράφεται σε tRNA θα εντοπίζεται η αλληλουχία 3’ACC5′. Τέτοια τράδα βάσεων εντοπίζεται μόνο στο Γ2, στην αλυσίδα ΙΙ, όπως αναγράφεται στο σχήμα.
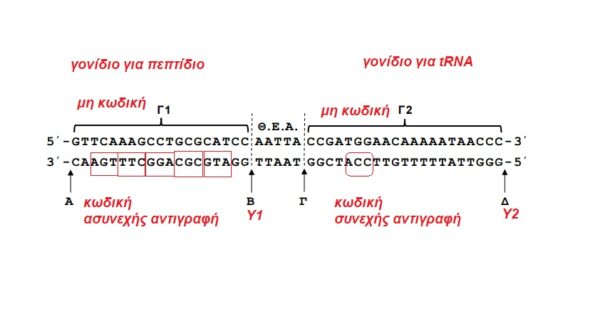
Αναγκαστικά, το Γ1 κωδικοποιεί το μικρό πεπτίδιο. Όσον αφορά τα γονίδια που μεταγράφονται σε mRNA και τελικά μετφράζονται σε πρωτείνες, ισχύει ότι στην κωδική τους αλυσίδα εντοπίζεται κωδικόνιο έναρξης (ΚΕ: 5’ATG3′), κωδικόνιο λήξης (ΚΛ: ένα από τα 5’TAA3′, 5’TAG3′, 5’TGA3′). Η ανάγνωση των βάσεων μεταξύ ΚΕ και ΚΛ γίνεται αν τρεις βάσεις (επειδή ο γενετικός κώδικας -ΓΚ είναι τριπλέτας), κατα συνεχή (επειδή ο ΓΚ είναι συνεχής) και μη επικαλυτπόμενο (επειδή ο ΓΚ είναι μη επικαλυπτόμενος) τρόπο.
Θεωρώντας ότι το Γ1 είναι συνεχές (δεν διαθέτει εσώνια), κωδική αλυσίδα του είναι η αλυσίδα ΙΙ, όπως αναγράφεται στο σχήμα.
ΣΗΜΕΙΩΣΗ: στην εκφώνηση έπρεπε να δίνεται ως δεδομένο ότι τα δυο γονίδια είναι συνεχή.
Γ2:
Η RNA πολυμεράση προσδένεται σε ειδικές περιοχές του DNA, που ονομάζονται υποκινητές. Οι υποκινητές βρίσκονται πάντοτε πριν από την αρχή κάθε γονιδίου. Κατά την έναρξη της μεταγραφής ενός γονιδίου η RNA πολυμεράση προσδένεται στον υποκινητή και προκαλεί τοπικό ξετύλιγμα της διπλής έλικας του DNA. Η RNA πολυμεράση συνδέει τα ριβονουκλεοτίδια που προστίθενται το ένα μετά το άλλο, με 3′-5’φωσφοδιεστερικό δεσμό. Η μεταγραφή έχει προσανατολισμό 5’→3′.
Με βάση τα παραπάνω, η θέση του υποκινητή για το Γ1 είναι το σημείο Β και για το Γ2 το Δ.
Γ3:
Η αντιγραφή του DNA αρχίζει από καθορισμένα σημεία, που ονομάζονται θέσεις έναρξης της αντιγραφής (Θ.Ε.Α.). Για να αρχίσει η αντιγραφή του DNA, είναι απαραίτητο να ξετυλιχθούν στις θέσεις έναρξης της αντιγραφής οι δύο αλυσίδες. Αυτό επιτυγχάνεται με τη βοήθεια ειδικών ενζύμων, που σπάζουν τους δεσμούς υδρογόνου μεταξύ των δύο αλυσίδων. Τα ένζυμα αυτά ονομάζονται DNA ελικάσες. Όταν ανοίξει η διπλή έλικα, δημιουργείται μια «θηλιά», η οποία αυξάνεται και προς τις δύο κατευθύνσεις. Τα κύρια ένζυμα που συμμετέχουν στην αντιγραφή του DNA ονομάζονται DNA πολυμεράσες. Επειδή τα ένζυμα αυτά δεν έχουν την ικανότητα να αρχίσουν την αντιγραφή, το κύτταρο έχει ένα ειδικό σύμπλοκο που αποτελείται από πολλά ένζυμα, το πριμόσωμα, το οποίο συνθέτει στις θέσεις έναρξης της αντιγραφής μικρά τμήματα RNA, συμπληρωματικά προς τις μητρικές αλυσίδες, τα οποία ονομάζονται πρωταρχικά τμήματα. DNA πολυμεράσες επιμηκύνουν τα πρωταρχικά τμήματα, τοποθετώντας συμπληρωματικά δεοξυριβονουκλεοτίδια απέναντι από τις μητρικές αλυσίδες του DNA. Τα νέα μόρια DNA αρχίζουν να σχηματίζονται, καθώς δημιουργούνται δεσμοί υδρογόνου μεταξύ των συμπληρωματικών αζωτούχων βάσεων των δεοξυριβονουκλεοτιδίων. Οι DNA πολυμεράσες λειτουργούν μόνο προς καθορισμένη κατεύθυνση και τοποθετούν τα νουκλεοτίδια στο ελεύθερο 3′ άκρο της δεοξυριβόζης του τελευταίου νουκλεοτιδίου κάθε αναπτυσσόμενης αλυσίδας. Έτσι, λέμε ότι αντιγραφή γίνεται με προσανατολισμό 5′ προς 3′. Κάθε νεοσυντιθέμενη αλυσίδα θα έχει προσανατολισμό 5’→3′. Έτσι, σε κάθε διπλή έλικα που παράγεται οι δύο αλυσίδες θα είναι αντιπαράλληλες. Για να ακολουθηθεί αυτός ο κανόνας σε κάθε τμήμα DNA που γίνεται η αντιγραφή, η σύνθεση του DNA είναι συνεχής στη μια αλυσίδα και ασυνεχής στην άλλη.
Κατ’ επέκταση για το Γ1 ισχύουν: μη κωδική – συνεχώς, κωδική – ασυνεχώς, ενώ για το Γ2 ισχύουν: κωδική – συνεχώς, μη κωδική – ασυνεχώς.
Γ4:
DNA βιβλιοθήκη ονομάζεται η συλλογή κλωνοποιημένων τμημάτων DNA που είτε αντιπροσωπεύουν ολόκληρο το γονιδίωμα (γονιδιωματική βιβλιοθήκη) ή αντιπροσωπεύουν DNA αντίγραφα του ολικού mRNA που παράγεται από ένα κύτταρο ή ιστό (cDNA βιβλιοθήκη).
Κατ’ επέκταση, στη γονιδιωματική βιβλιοθήκη μπορούν να κλωνοποιηθούν για να μελετηθούν όλα τα γονίδια που εντοπίζονται στο γονιδίωμα, υπό τηνπρουπόθεση η χρησιμοποιούμενη περιοριστική ενδονουκλεάση να δρα εκατέρωθεν τους.
Σε cDNA βιβλιοθήκη μπορούν να κλωνοποιηθούν μόνο τα ενεργά (λόγω γονιδιακής ρύθμισης) γονίδια που μεταγράφονται σε mRNA. Άρα δεν μπορούν να κλωνοποιηθούν τα γονίδια που μεταγράφονται σε tRNA, rRNA, snRNA. Επιπλέον, σε cDNA βιβλιοθήκες ευκαρυωτικών κυττάρων, κλωνοποιούνται μόνο οι αλληλουχίες που αντιστοιχούν στο ώριμο mRNA, δηλαδή τα εξώνια (στα οποία συμπεριλαμβάνονται η κωδικοποιούσα περιοχή – κωδικόνια και οι αμετάφραστες περιοχές).
Με βάση τα παραπάνω, το γονίδιο που μεταγράφεται σε tRNA μπορεί να κλωνοποιηθεί μόνο με γονιδιωματική βιβλιοθήκη.
Συνεχές γονίδιο που μεταγράφεται σε mRNA μπορεί να κλωνοποιηθεί τόσο με γονιδιωματική όσο και με cDNA βιβλιοθήκη. Για την παραγωγή πεπτιδίου σε μεγάλες ποσότητες, πέραν της κλωνοποίησης του γονιδίου, απαραίτητη προυπόθεση είναι η έκφραση του, γεγονός που επιτυγχάνεται εφόσον ο φορέας κλωνοποίησης (ΦΚ) περιέχει κατάλληλο υποκινητή που θα αναγνωρίζεται από το συνδυασμό μεταγραφικων παραγόντων του ξενιστή που θα μετασχηματιστεί. Η φορά ανασυνδυασμού του γονιδίου στον ΦΚ πρέπει να έχει ως αποτέλεσμα η κωδική αλυσίδα του υπό κλωνοποίσηση γονιδίου να είναι τμήμα της κωδικής αλυσίδας του μεταγραφόμενου τμήματος του ΦΚ.
Με βάση τα παραπάνω, το γονίδιο για το πεπτίδιο μπορεί να κλωνοποιηθεί και με τα δυο είδη DNA βιβλιοθήκης.
Γ5:
α) όπως φαίνεται στο σχήμα
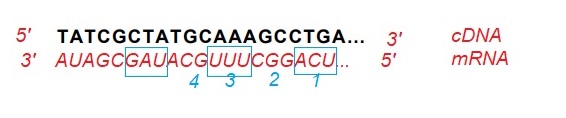
β)
ΕΚΔΟΧΗ 1
υπάρχουν δυο περιπτώσεις
1η περίπτωση: το τμήμα του mRNA που αναγράφεται ανήκει εξολοκληρου στην 3′ αμετάφραστη περιοχή. Σε αυτήν την περίπτωση, δεν κωδικοποιείται κανένα αμινοξύ
2η περίπτωση: το τμήμα του mRNA που αναγράφεται περιέχει το κωδικόνιο λήξης 5’UAG3′. Σε αυτήν την περίπτωση ο αριθμός των αμινοξέων που κωδικοποιούνται είναι 4.
ΕΚΔΟΧΗ 2
Το cDNA αποτελεί τμήμα της μη κωδικής αλυσίδας ασυνεχούς γονιδίου. Η μη κωδική αλυσίδα γονιδίου δεν κωδικοποιεί αμινοξέα. Αμινοξέα κωδικοποιούνται από το mRNA (συγκεκριμένα από την κωδικοποιούσα περιοχή του). Επειδή, ο όρος κωδικόνιο χρησιμοποιείται και στην κωδική αλυσίδα γονίδιου που μεταγράφεται σε mRNA, καθ’ υπερβαση μπορούμε να πούμε πως η κωδικη αλυσίδα κωδικοποιεί αμινοξέα. Δεν ισχύει όμως κάτι τέτοιο για την μη κωδική.
Η σωστή λοιπόν απάντηση είναι: δεν κωδικοποιεί αμινοξύ.
Σ.Σ.: ο θεματοδότης, πιθανότατα σκέφτεται την πρώτη εκδοχή, 2η περίπτωση (την οποία διάβασε σε γνωστό βοήθημα, γνωστού φροντιστηρίου). Ο έξυπνος υποψήφιος πρέπει να γράψει και τις δυο εκδοχές.
ΘΕΜΑ Δ
Δ1
Όσον αφορά το γονίδιο για το μεταγραφικό παράγοντα (ΜΠ) Δ: είναι αυτόσωμικό καθώς εντοπίζεται στο 4ο ζεύγος ομολόγων χρωμοσωμάτων. Με βάση την εκφώνηση πιθανολογούμε ότι υπαρχουν δυο αλληλόμορφά του:
Δ: παραγωγή ΜΠ (επικρατές)
δ: μη παραγωγή ΜΠ (υπολοιπόμενο)
πιθανοί γονότυποι – φαινότυποι: ΔΔ (ομόζυγος) Δδ (ετερόζυγος, δδ (ομόζυγος)
Όσον αφορά το γονίδιο που ελέγχει τη σύνθεση του αντιαιμορροφιλικού παράγοντα VIII: είναι φυλοσύνδετο καθώς εντοπίζεται εντοπίζεται στη φυλοζύνδετη περιοχή του χρωμοσώματος Χ.
ΧΑ: παραγωγή παράγοντα VIII (επικρατές)
Xα: μη παραγωγή παράγοντα VIII (υπολειπόμενο)
πιθανοί γονότυποι – φαινότυποι
ΧΑΧΑ: θηλυκό που παράγει τον παράγοντα VIII, ΧΑΧα : θηλυκό που παράγει τον παράγοντα VIII – φορέας, ΧαΧα: θηλυκό που δεν παράγει τον παράγοντα VIII (αιμορροφιλία Α)
ΧΑΨ: αρσενικό που παράγει τον παράγοντα VIII, XαΨ: αρσενικό που δεν παράγει τον παράγοντα VIII (αιμορροφιλία Α)
Τα δυο γονίδια είναι ανεξάρτητα, καθώς εντοπίζονται σε διαφορετικά ζεύγη χρωμοσωμάτων.
Πιθανοί γονότυποι διυβριδισμού – πιθανοί φαινότυποι:
ΔΔΧΑΧΑ : θηλυκό που παράγει τον παράγοντα VIII
ΔΔΧΑΧα : θηλυκό που παράγει τον παράγοντα VIII
ΔΔΧαΧα: θηλυκό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
ΔΔΧΑΨ: αρσενικό που παράγει τον παράγοντα VIII
ΔΔΧαΨ: αρσενικό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
ΔδΧΑΧΑ : θηλυκό που παράγει τον παράγοντα VIII
ΔδΧΑΧα : θηλυκό που παράγει τον παράγοντα VIII
ΔδΧαΧα: θηλυκό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
ΔδΧΑΨ: αρσενικό που παράγει τον παράγοντα VIII
ΔδΧαΨ: αρσενικό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
δδΧΑΧΑ : θηλυκό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
δδΧΑΧα : θηλυκό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
δδΧαΧα: θηλυκό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
δδΧΑΨ: αρσενικό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
δδΧαΨ: αρσενικό που δεν παράγει τον παράγοντα VIII και έχει αιμορροφιλία Α
Ο γονότυπος δδ δεν επιτρέπει την έκφραση του γονιδίου γεγονός που οδηγεί σε αιμορροφίλία Α ανεξάρτητα του γονοτύπου για το γονίδιο της αντιαιμορροφιλικής πρωτεινης.
Η διαστάυρωση είναι ΔδΧΑΨ (πατέρας) * ΔδΧΑΧα (μητέρα).
Τα αποτελέσματα της φαίνονται στο τετράγωνο του Punnett:
| ΔΧΑ | δΧΑ | ΔΨ | δΨ | |
| ΔΧΑ | ΔΔΧΑΧΑ | ΔδΧΑΧΑ | ΔΔΧΑΨ | ΔδΧΑΨ |
| ΔΧα | ΔΔΧΑΧα | ΔδΧΑΧα | ΔΔΧαΨ | ΔδΧαΨ |
| δΧΑ | ΔδΧΑΧΑ | δδΧΑΧΑ | ΔδΧΑΨ | δδΧΑΨ |
| δΧα | ΔδΧΑΧα | δδΧΑΧα | ΔδΧαΨ | δδΧαΨ |
Φαινοτυπική αναλογία:
6 θηλυκό χωρίς αιμορροφιλία : 2 θηλυκό με αιμορροφιλία Α: 3 αρσενικό χωρίς αιμορροφιλία : 5 αρσενικό με αιμορροφιλία Α
Δ2
α) ΕΚΔΟΧΗ 1
τα σωματικά κύτταρα που παράγουν και εκκρίνουν στο αίμα τον παράγοντα VIII ανήκουν στο αιμοποιητικό σύστημα, οπότε με βάση το σχολικό βιβλίο: Τα κύτταρα του αιμοποιητικού συστήματος μπορούν να τροποποιούνται γενετικά, να αναπτύσσονται σε κυτταροκαλλιέργειες και να εισάγονται με ενδοφλέβια ένεση στον οργανισμό. Ο τύπος αυτός της γονιδιακής θεραπείας ονομάζεται ex vivo, γιατί τα κύτταρα τροποποιούνται έξω από τον οργανισμό και εισάγονται πάλι σ’ αυτόν.
ΕΚΔΟΧΗ 2
τα σωματικά κύτταρα που παράγουν και εκκρίνουν στο αίμα τον παράγοντα VIII ΔΕΝ ανήκουν στο αιμοποιητικό σύστημα, οπότε πάλι με βάση το σχολικό βιβλίο (τροποποιημένο): Τι γίνεται όμως αν πρέπει να τροποποιηθούν κύτταρα ενός άλλου οργάνου; Ο Anderson και οι συνεργάτες του πρότειναν μια άλλη προσέγγιση. Ανέπτυξαν «έξυπνους» φορείς, οι οποίοι προσβάλλουν τα κύτταρα του ιστού που πάσχει. Συγκεκριμένα, τα φυσιολογικά γονίδια ενσωματώνονται σε μόρια-φορείς, που εισάγονται κατευθείαν στον οργανισμό. Το είδος αυτό της γονιδιακής θεραπείας ονομάζεται in vivo. Το φυσιολογικό γονίδιο ενσωματώνεται αρχικά σε έναν αδενοϊό. Ο ανασυνδυασμένος ιός εισέρχεται στον οργανισμό με ψεκασμό και μόλυνει τα κύτταρα του αντίστοιχου οργάνου. Μετά την εισαγωγή του στα κύτταρα, το φυσιολογικό γονίδιο ενσωματώνεται στο γονιδίωμά τους και μπορεί να παράγει το φυσιολογικό προϊόν.
Σ.Σ.: ο παράγοντας VIII παράγεται και εκκρίνεται από το ήπαρ. Ο σωστός τύπος γονιδιακής θεραπείας είναι in vivo. Όμως η γνώση του οργάνου παραγωγής του συγκεκριμένου παράγοντα είναι εκτός ύλης. Κατά συνέπεια ο υποψήφιος δεν μπορεί να προσεγγίσει τη σωστή απάντηση. Ο θεματοδότης πιθανότατα σκέφτεται την πρώτη εκδοχή (την οποία διάβασε στο ίδιο βοήθημα). Ο έξυπνος υποψήφιος πρέπει να γράψει και τις δυο εκδοχές.
β)
Η RNA πολυμεράση προσδένεται στους υποκινητές, με τη βοήθεια πρωτεϊνών που ονομάζονται μεταγραφικοί παράγοντες. Οι υποκινητές και οι μεταγραφικοί παράγοντες αποτελούν τα ρυθμιστικά στοιχεία της μεταγραφής του DNA και επιτρέπουν στην RNA πολυμεράση να αρχίσει σωστά τη μεταγραφή. Στους ευκαρυωτικούς οργανισμούς οι μεταγραφικοί παράγοντες παρουσιάζουν τεράστια ποικιλία. Κάθε κυτταρικός τύπος περιέχει διαφορετικά είδη μεταγραφικών παραγόντων. Διαφορετικός συνδυασμός μεταγραφικών παραγόντων ρυθμίζει τη μεταγραφή κάθε γονιδίου. Μόνο όταν ο σωστός συνδυασμός των μεταγραφικών παραγόντων προσδεθεί στον υποκινητή ενός γονιδίου, αρχίζει η RNA πολυμεράση τη μεταγραφή ενός γονιδίου.
Κατ’ επέκταση, η ενσωμάτωση του ΧΑ στους γονότυπους δδΧΑΧΑ , δδΧΑΧα, δδΧαΧα, δδΧΑΨ, δδΧαΨ, είτε γίνει in vivo είτε ex vivo, δεν θα έχει πρακτικό αποτέλεσμα γιατί απουσιάζει ο μεταγραφικός παράγοντας Δ από τα κύτταρα του ασθενούς που τροποποιούνται:
Κατά συνέπεια, η γονιδιακή θεραπεία θα έχει αποτέλεσμα μόνο στους γονότυπους ΔΔΧαΨ και ΔδΧαΨ
Δ3
έστω 3Α: τροποποιημένο χρωμόσωμα 3 που περιέχει το γονίδιο για την ανθρώπινη α1 – αντιθρυψίνη
5Α: τροποποιημένο χρωμόσωμα 5 που περιέχει το γονίδιο για την ανθρώπινη α1 – αντιθρυψίνη
Οι γονότυποι των προβάτων που διασταυρώνονται είναι: 33Α55 (αρσενικό) * 3355Α(θηλυκό)
| 35 | 35Α | |
| 35 | 3355 (μη διαγονιδιακό) | 3355Α (διαγονιδιακό) |
| 3Α5 | 33Α55 (διαγονιδιακό) | 33Α55Α (διαγονιδιακό) |
η πιθανότητα (P) γέννησης διαγονιδιακού προβάτου είναι 3/4
για την παραγωγή γάλατος, πρέπει ο απόγονος να είναι θηλυκός : P(θηλυκός απόγονος) = 1/2
τα δυο γεγονότα (γέννηση διαγονιδιακού απογόνου – γέννηση θηλυκού απογόνου) κληρονομούνται ανεξάρτητα (3Α και 5Α είναι αυτοσωμικά γονίδια, ενώ το φύλο καθορίζεται από τα φυλετικά χρωμοσώματα). Η συνολική πιθανότητα είναι P (θηλυκού διαγονιδιακού απογόνου) = P (διαγονιδιακού απογόνου) * P (θηλυκού απογόνου) = 3/4* 1/2 = 3/8 (37,5%)
Δ4
έστω: 12Α το αλληλόμορφο για την παραγωγή του ενζύμου, που εντοπίζεται στο 12 φυσιολογικό χρωμόσωμα
12α το αλληλόμορφο για την μη παραγωγή του ενζύμου, που εντοπίζεται στο 12 φυσιολογικό χρωμόσωμα
12ε το μεταλλαγμένο χρωμόσωμα 12, με την έλλειψη της συγκεκριμένης γενετικής θέσης
πιθανοί γονότυποι – φαινότυποι – καρυότυποι
12Α 12Α : παραγωγή ενζύμου, καρυότυπος κατά φύσιν (ΚΦ)
12Α 12α : παραγωγή ενζύμου, καρυότυπος ΚΦ
12α 12α : μη παραγωγή ενζύμου, καρυότυπος ΚΦ
12Α 12ε : παραγωγή ενζύμου, καρυότυπος με έλλειψη τμήματος ενός χρωμοσώματος 12 (ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12)
12α 12ε : μη παραγωγή ενζύμου, καρυότυπος με ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12
12ε 12ε : μη παραγωγή ενζύμου, καρυότυπος με έλλειψη τμήματος από τα ομόλογα χρωμοσώματα 12 (ομόζυγη έλλειψη τμήματος χρωμοσώματος 12)
12α 12ε Χ Ψ * 12Α 12α Χ Χ
| 12Α Χ | 12α Χ | |
| 12α Χ | 12Α12α Χ Χ | 12α12α Χ Χ |
| 12ε Χ | 12Α 12ε Χ Χ | 12α12ε ΧΧ |
| 12α Ψ | 12Α12α Χ Ψ | 12α12α ΧΨ |
| 12ε Ψ | 12Α12ε ΧΨ | 12α12ε ΧΨ |
Φαινοτυπική αναλογία:
1 θηλυκό που παράγει το ένζυμο και ΚΦ καρυότυπο : 1 θηλυκό που δεν παράγει το ένζυμο και ΚΦ καρυότυπο : 1 θηλυκό που παράγει το ένζυμο και καρυότυπο με ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12 : 1 θηλυκό που δεν παράγει το ένζυμο και καρυότυπο με ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12 : 1 αρσενικό που παράγει το ένζυμο και ΚΦ καρυότυπο : 1 αρσενικό που δεν παράγει το ένζυμο και ΚΦ καρυότυπο : 1 αρσενικό που παράγει το ένζυμο και καρυότυπο με ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12 : 1 αρσενικό που δεν παράγει το ένζυμο και καρυότυπο με ετερόζυγη έλλειψη τμήματος χρωμοσώματος 12
α) P(12α 12α Χ Ψ ή 12α 12ε Χ Ψ) = 2/8 (25%)
β) P (12α 12ε Χ Ψ σε θηλυκό απόγονο) = 0 % καθώς η παρουσία του χρωμοσώματος Ψ προσδιορίζει ως αρσενικό το φύλο του απογόνου.
Comments are closed.

Τα θέμα και φέτος είχαν ασάφειες σε κάποιες εκφωνήσεις. Ορισμένα θέματα προέρχονται από συγκεκριμένο βοήθημα το οποίο δυστυχώς βρίθει από λάθη, και οι θεματοδότες πιθανότατα δεν μπορούν να εντοπίσουν.